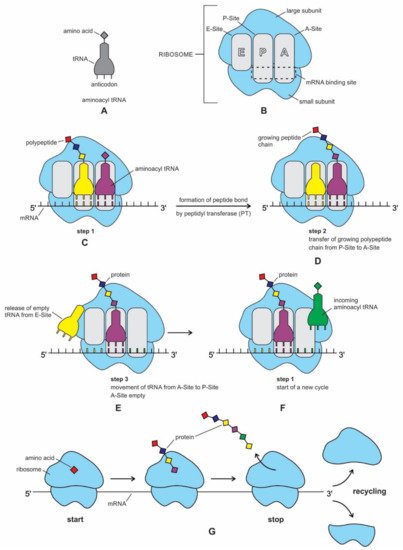
Es gibt mal wieder einen neuen
Überblicksartikel zum eigentlich relevanten Problem der Abiogenese, nämlich der Entstehung des Translationsmechanismus und des genetischen Codes. Erschienen ist er am 1. März diesen Jahres in der Online-Zeitschrift "Life", die in Basel verlegt wird und eine einschlägig mit der Abiogense befasste Fachzeitschrift ist, in der Akademiker dem Publikationszwang nachgeben dürfen, dem sie in ihrer Karriere ausgesetzt sind.
Die aktuelle Arbeit ist in 10 Abschnitte untergliedert, von denen neben der Einleitung und der Zusammenfassung insbesondere die Abschnitte 5., 6. und 7. für dieses Thema von Interesse sind:
1. Introduction
2. Peptide/RNA World
3. The Age of Information
4. The Use of Information Theory in Biology
5. Temporal Order of Emergence of the Translation Machines
6. The Origin and Evolution of the Genetic Code
7. Coevolution of Translation Machines and the Genetic Code
8. Design of Translation Machines and the Genetic Code
9. Simulation and Visualization of the Translation Pathways
10. Discussion and Conclusions
Das Vorgeplänkel zur Peptid/RNA-Welt sowie zum Zeitalter der Information und der Anwendung der Informationstheorie in der Biologie übergehe ich an dieser Stelle komplett, da sie nur den heutigen Ist-Stand der Molekularbiologie reflektieren. Das Nachgeplänkel zum Design von Translationsmaschinen und dem Genetischen Code sowie der Simulation und Visualisierung der Translations-Entwicklungswege lasse ich ebenfalls unberücksichtigt, da ich von solchen Rechenspielereien nicht viel halte.
Interessant ist bei diesem Artikel aber die Darlegung des aktuellen Diskurses über verschiedene Modelle, die mit der Entstehung der Translation und des genetischen Codes in Verbindung stehen. In dieser Übersichtlichkeit finden sich solche Darstellungen nicht oft, so dass ich mich daher auf die genannten drei Abschnitte konzentrieren werde. Der Umfang des Artikels von immerhin 73 Seiten im pdf ist recht groß, so dass man diesem Artikel doch eine recht große Menge an einschlägigen Informationen entnehmen kann.
Hilfreich sind einige Illustrationen, die dem Text beigefügt sind sowie die Erläuterungen zu diesen Illustrationen. Ich werde mich daher vorrangig mit diesen Illustrationen befassen und die beigefügten Erläuterungen als Übersetzung kommentieren, um diesen Beitrag nicht zu lang werden zu lassen.
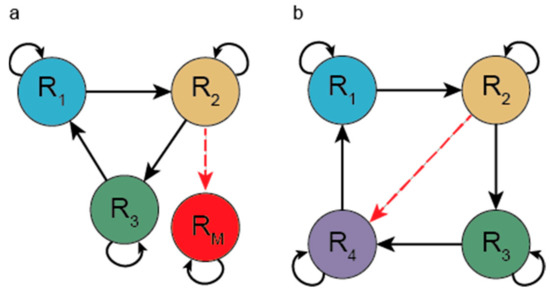
Zunächst wird am Ende des Einleitungstextes von Abschnitt 5. hervorgehoben, dass man 9 Stadien der zeitlichen Abfolge der Entstehung der Translation identifiziert hat:
Wir identifizieren neun Hauptstufen für die Entstehung und Entwicklung des Translationsmaschinenkomplexes und des genetischen Codes, der zur Proteinsynthese führt. Diese möglichen biochemischen Wege sind:
(1) die Auswahl von Aminosäuren;
(2) der Ursprung der RNA;
(3) der Ursprung des Ribozyms;
(4) der Ursprung der Transfer-RNA;
(5) den Ursprung des Stoffwechsels;
(6) der Ursprung der Aminoacyl-tRNA-Synthetase;
(7) der Ursprung der Messenger-RNA und der Übersetzung;
(8) der Ursprung des Ribosoms; und schließlich
(9) Proteinsynthese.
Während der Entstehung dieser biochemischen Pfade kooperierten der genetische Code und das Übersetzungssystem mit der Übersetzungsmaschine.
Diese neun Hauptstufen werden in der Folge der Reihe nach abgehandelt. Die Auswahl der Aminosäuren sei hierbei in zwei Stadien erfolgt: Zunächst die vier am einfachsten in Miller-Urey-Reaktionen entstehenden Aminosäuren (Glycin, Alanin, Valin, Asparaginsäure), später dann weitere sechs (Glutaminsäure, Leucin, Prolin, Histidin, Arginin und Glutamin). Hierbei sollen von Anfang an Vorläufer der heutigen tRNA's und der heutigen aaRS' beteiligt gewesen sein, die in Form von kürzeren RNA-Strängen sowie kürzeren Peptiden vorhanden gewesen sein sollen. Details dazu werden später ausgeführt.
Die Entstehung und Entwicklung von RNA wird in sechs hypothetischen Stufen dargestellt:
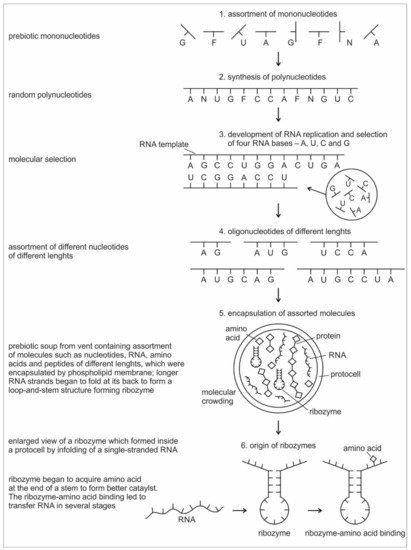
Diese Darstellung ist von unten nach oben zu lesen. Die Bildunterschrift listet die sechs Stadien auf:
Abbildung 2. Sechs Hauptschritte repräsentieren die frühe Entwicklung von nicht kodierender RNA in der hydrothermalen Schlot-Umgebung.
In der ersten Stufe gibt es eine Auswahl verschiedener Nukleotide (einschließlich einiger, die nicht in RNA enthalten sind).
In der zweiten Stufe werden diese Nukleotide durch Polymerisation unter Entfernung von Wassermolekülen statistisch zufällig zu Polynukleotiden zusammengefügt.
In der dritten Stufe wurden die vier Nukleotide A, U, G und C während der Replikation durch die Watson-Crick-Basenpaarung ausgewählt.
In der vierten Stufe durchlaufen die Nukleotide eine Polymerisation, um eine Mischung von Polynukleotiden zu erzeugen, die in Länge und Reihenfolge zufällig sind.
In der fünften Stufe werden eine Vielzahl von Biomolekülen aus der Schlot-Umgebung, wie Aminosäuren, Mononukleotide, Oligonukleotide und Peptide, willkürlich eingekapselt, wodurch ein molekulares Gedränge entsteht. Aufgrund der Überfüllung beginnt sich die einzelsträngige RNA zu falten und bildet den doppelsträngigen Stamm und die einzelsträngige Schleife, aus der sich die Haarnadel bildet.
In der sechsten Stufe wird diese Sekundärstruktur der RNA separat gezeigt: Sie bildet ein Ribozym und beginnt, als Enzym zu wirken. Stämme entstehen durch Wasserstoffbrücken zwischen komplementären Basenpaaren. Das Ribozym erhält Aminosäuren an der CCA-Sequenz des Stammes als "Cofaktoren", die seine katalytische Effizienz erhöhen. Das gegenüberliegende Ende der Schleife besteht aus drei ungepaarten Basen, die nach außen weisen und eine Bindungsstelle für die Anbringung von drei entsprechenden Mononukleotiden bilden. Dies ist der Beginn des Auftauchens der Proto-tRNA.
Fraglich ist insbesondere die sechste Stufe, wo ausgeführt wird, dass sich die Haarnadelschleife der proto-tRNA so ausbildet, dass sich drei freie ungepaarte Basen an einem Ende der Schleife ausbilden. Es wird nicht dargelegt, warum das erfolgen sollte. Auch der Artikeltext gibt hierzu nichts her. Denkbar ist, dass die eine Seite der Haarnadelschleife mit einer Aminosäure belegt ist, so dass sich die gegenständigen drei Basen des komplementären Stranges nicht mehr an das andere Ende anlagern können. Aber dann taucht die Frage auf, warum das andere Ende nicht ebenfalls mit einer Aminosäure als Kofaktor belegt wird. An dieser Stelle wird es also brüchig in der Argumentation.
Aus der Bildung von Haarnadelschleifen leitet sich nicht nur die Entstehung von proto-tRNA's ab, sondern darüber hinaus auch die Entstehung von Ribozymen - also RNA-Molekülen, deren Struktur dazu geeignet ist, enzymatische Funktionen zu erfüllen, die für die Regulierung von Stoffwechselprozessen bedeutsam ist. Einige dieser Ribozyme sollen dann auch in der Lage sein, Aminosäuren zu Peptiden zu verketten. Auch hierzu wird eine Abbildung beigefügt:
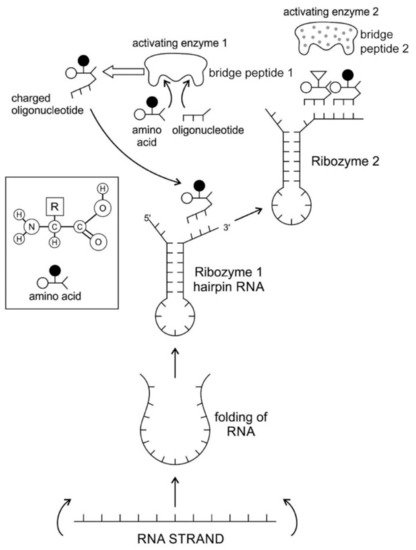
Wiederum ist diese Abbildung von unten nach oben zu lesen. Der Begleittext führt aus:
Abbildung 3. Der Ursprung von Haarnadel-Ribozym und seine chemische Bindung mit einer geeigneten Aminosäure.
Eine einzelsträngige RNA kann eine Sekundärstruktur entwickeln, indem sie mit einem doppelsträngigen Stamm und einer einzelsträngigen Schleife ein Haarnadel-Ribozym bildet.
Das Ribozym erhielt Aminosäure als Cofaktor, um einen effizienteren Katalysator zu bilden.
Die Aminosäure wird an ein Oligonukleotid (ein RNA-Molekül, das nur drei Nukleotide enthält) durch ein Aktivierungsenzym wie ein "Brückenpeptid" gebunden, und das Oligonukleotid wird durch Basenpaarung (Ribozym 1) an die Oberfläche des Ribozyms gebunden.
Das aktivierende Enzym 2 würde die nächste geladene Aminosäure binden, und das Oligonukleotid wird an Ribozym 2 gebunden, wobei die Peptidbindung gebildet wird.
Es ist hierbei wieder fraglich, warum sich Aminosäuren nur an dreibasige Oligonucleotide binden und nicht an zweibasige oder vierbasige. Weiterhin ist fraglich, warum nur dreibasige Oligonucleotide an die entsprechenden Ribozyme andocken und nicht zweibasige oder vierbasige. Hier wird bereits eine Entwicklung vorweggenommen, die zum späteren Triplett-Code führt, allerdings ohne auf die Selektionsdrücke einzugehen, die zur Favorisierung der dreibasigen Variante führen.
Die fraglichen Selektionsdrücke müssten jenseits der molekularen Grundlage angesiedelt sein, da die freiliegenden sechs Basen des Ribozyms sowohl zwei dreibasige Oligonucleotide andocken ließen wie auch drei zweibasige sowie eine zweibasige und eine vierbasige Variante. Hierzu wird nichts im Text angeführt, so dass auch hier eine gewisse Brüchigkeit in der Argumentation vorliegt.
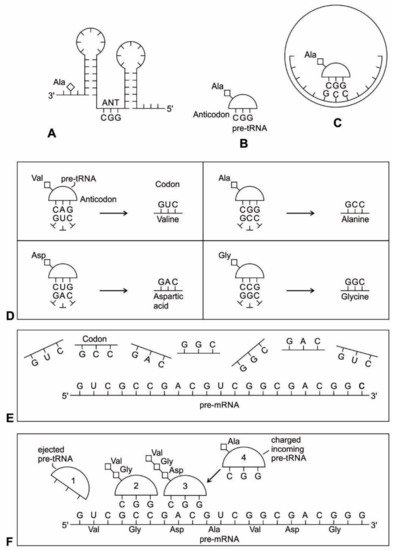
Zur Entstehung der tRNA ist diese Abbildung beigefügt:
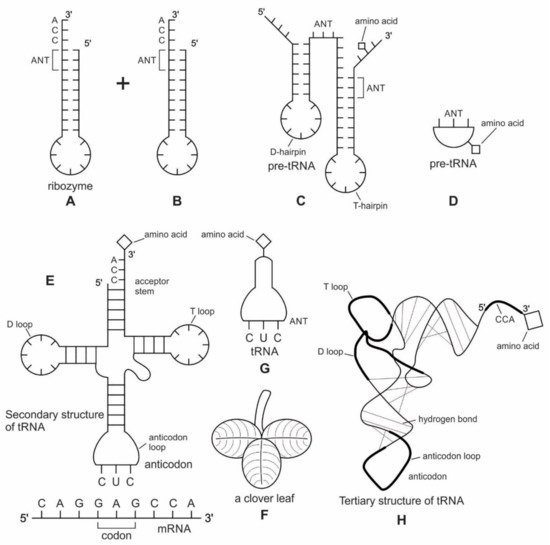
Die Bildunterschrift lautet:
Abbildung 4: Das Doppelhaarnadelmodell der Transfer-RNAs (tRNA) -Bildung zeigt die evolutionären Übergänge.
(A, B) zeigt eine sekundäre Haarnadelstruktur von zwei RNA-Molekülen (wie z. B. Ribozymen) mit jeweils einem Stamm und einer Schleife: Die CCA-Sequenz am 3'-Ende des Stamms bietet eine Bindungsstelle für eine Aminosäure, während das 5'-Ende bietet eine Bindungsstelle für Phosphor;
(C) kann die direkte Verdoppelung oder Ligation der Haarnadelstruktur eine Doppelhaarnadelstruktur erzeugt haben, die eine D-Haarnadel und eine T-Haarnadel erzeugt. Eine Anticodon (ANT) -Stelle bildet sich zwischen den beiden Stielen. In diesem neu konfigurierten Prä-tRNA-Molekül befinden sich die Akzeptor- und die Anticodon-Stelle jetzt näher zusammen, sodass ein Prä-Messenger-RNA (mRNA) -Molekül für die Proteinsynthese entschlüsselt werden kann.
(D) ein schematisches Diagramm, das die hervorstechenden Merkmale des Prä-tRNA-Moleküls mit der Anticodon-Stelle zeigt;
(E) könnte das zeitgenössische tRNA-Molekül voller Länge durch die Ligation von zwei halben prä-tRNA-Strukturen gebildet worden sein. Seine Akzeptorstammbasen und Anticodonstamm- / Schleifenbasen an der tRNA-5'-Hälfte und der 3'-Hälfte passen zur Doppelhaarnadel-Faltung. Dies lässt vermuten, dass sich die ursprünglichen Doppelhaarnadel-RNA-Moleküle zur modernen tRNA entwickelt haben könnten. Diese neue Sekundärstruktur der tRNA ähnelt einem Kleeblatt. Ihr Anticodon-Ende bildet ein komplementäres Basenpaar mit dem mRNA-Codon.
(F), ein Kleeblatt aus der Natur, veranschaulicht die strukturelle Ähnlichkeit mit dem neuen tRNA-Molekül;
(G), ein schematisches Diagramm, das die hervorstechenden Merkmale des tRNA-Moleküls zeigt, wobei das Anticodon hervorgehoben wird. Die tRNA spielt eine entscheidende Rolle beim Abgleichen einer Aminosäure mit einem spezifischen Codon. Wenn tRNA an eine Aminosäure gebunden ist, spricht man von Aminoacyl-tRNA. Für jede Aminosäure gibt es jetzt eine entsprechende tRNA mit einem geeigneten Anticodon;
(H), faltet sich die Kleeblatt-Sekundärstruktur der tRNA zur L-förmigen Tertiärstruktur. Am CCA-Minihelix-Ende interagiert die Aminoacylierungsstelle mit einer großen ribosomalen Einheit, um eine Peptidbindung zu bilden. Das entgegengesetzte Ende interagiert mit der kleinen ribosomalen Untereinheit, um mRNA-Tripletts durch Codon-Anticodon-Wechselwirkungen zu decodieren.
Die Vermutung, die heutige Kleeblatt-Sekundärstruktur der tRNA könnte auf eine Verbindung von zwei Haarnadelschleifen zurückzuführen sein, beruht auf Massimo DiGiulio aus dem Jahr 1992. Die Autoren dieses Artikels greifen dieses Modell auf und führen es hier im Rahmen ihres umfassenderen Szenarios mit an.
Zur Entstehung und Evolution des Stoffwechsels führen die Autoren an, dass es einen engen Bezug zum mineralischen Untergrund gegeben haben muss, der über die vorhandenen Metall-Ionen sowohl selektierend wie auch katalysierend auf die auf ihm ablaufenden Prozesse gewirkt hat. Insbesondere die Effekte von Zinksulfid in Tonmineralien werden hier näher erörtert:
Tonmineralien, die als Schwämme wirken, können Wasser und polare organische Moleküle zurückhalten. Sie könnten eine Schlüsselrolle bei der Konzentration und Katalyse der Polymerisation wichtiger organischer Moleküle wie RNA und Protein gespielt haben.
Später mehr zu den weiteren Schritten ...

JacobMonod schrieb:Er operiert zwar an einigen Stellen mit einschlägigen Formulierungen, die Du auch zitiert hattest, aber der Fokus liegt bei Monod m.E. auf der Unableitbarkeit aus den Grundlagen heraus, die - da diese sich aus der Zufälligkeit ergibt - den Schluss auf die nicht gegebene Notwendigkeit (im Sinne einer naturgesetzlichen Zwangsläufigkeit) erlaubt. Ob einzigartig oder nicht, bleibt schlicht unentscheidbar, da nicht ableitbar.